ID:AGO2_HUMAN DESCRIPTION: RecName: Full=Protein argonaute-2; Short=Argonaute2; Short=hAgo2; EC=3.1.26.n2; AltName: Full=Eukaryotic translation initiation factor 2C 2; Short=eIF-2C 2; Short=eIF2C 2; AltName: Full=PAZ Piwi domain protein; Short=PPD; AltName: Full=Protein slicer; FUNCTION: Required for RNA-mediated gene silencing (RNAi) by the RNA-induced silencing complex (RISC). The 'minimal RISC' appears to include EIF2C2/AGO2 bound to a short guide RNA such as a microRNA (miRNA) or short interfering RNA (siRNA). These guide RNAs direct RISC to complementary mRNAs that are targets for RISC- mediated gene silencing. The precise mechanism of gene silencing depends on the degree of complementarity between the miRNA or siRNA and its target. Binding of RISC to a perfectly complementary mRNA generally results in silencing due to endonucleolytic cleavage of the mRNA specifically by EIF2C2/AGO2. Binding of RISC to a partially complementary mRNA results in silencing through inhibition of translation, and this is independent of endonuclease activity. May inhibit translation initiation by binding to the 7- methylguanosine cap, thereby preventing the recruitment of the translation initiation factor eIF4-E. May also inhibit translation initiation via interaction with EIF6, which itself binds to the 60S ribosomal subunit and prevents its association with the 40S ribosomal subunit. The inhibition of translational initiation leads to the accumulation of the affected mRNA in cytoplasmic processing bodies (P-bodies), where mRNA degradation may subsequently occur. In some cases RISC-mediated translational repression is also observed for miRNAs that perfectly match the 3' untranslated region (3'-UTR). Can also up-regulate the translation of specific mRNAs under certain growth conditions. Binds to the AU element of the 3'-UTR of the TNF (TNF-alpha) mRNA and up-regulates translation under conditions of serum starvation. Also required for transcriptional gene silencing (TGS), in which short RNAs known as antigene RNAs or agRNAs direct the transcriptional repression of complementary promoter regions. CATALYTIC ACTIVITY: Endonucleolytic cleavage to 5'- phosphomonoester. ENZYME REGULATION: Inhibited by EDTA. BIOPHYSICOCHEMICAL PROPERTIES: Kinetic parameters: KM=1.1 nM for a synthetic 21-nucleotide single-stranded RNA; SUBUNIT: Interacts with DICER1 through its Piwi domain and with TARBP2 during assembly of the RNA-induced silencing complex (RISC). Together, DICER1, EIF2C2/AGO2 and TARBP2 constitute the trimeric RISC loading complex (RLC), or micro-RNA (miRNA) loading complex (miRLC). Within the RLC/miRLC, DICER1 and TARBP2 are required to process precursor miRNAs (pre-miRNAs) to mature miRNAs and then load them onto EIF2C2/AGO2. EIF2C2/AGO2 bound to the mature miRNA constitutes the minimal RISC and may subsequently dissociate from DICER1 and TARBP2. Note however that the term RISC has also been used to describe the trimeric RLC/miRLC. The formation of RISC complexes containing siRNAs rather than miRNAs appears to occur independently of DICER1. Interacts with EIF2C1/AGO1. Also interacts with DDB1, DDX5, DDX6, DDX20, DHX30, DHX36, DDX47, DHX9, EIF6, ELAVL, FXR1, GEMIN4, HNRNPF, IGF2BP1, ILF3, IMP8, MATR3, MOV10, PABPC1, PRMT5, P4HA1, P4HB, RBM4, SART3, TNRC6A, TNRC6B, UPF1 and YBX1. Interacts with the P-body components DCP1A and XRN1. Associates with polysomes and messenger ribonucleoproteins (mNRPs). Interacts with RBM4; the interaction is modulated under stress-induced conditions, occurs under both cell proliferation and differentiation conditions and in a RNA- and phosphorylation-independent manner. Interacts with LIMD1, WTIP and AJUBA. Interacts with TRIM71. INTERACTION: Q96C10:DHX58; NbExp=2; IntAct=EBI-528269, EBI-744193; Q9UPY3:DICER1; NbExp=6; IntAct=EBI-528269, EBI-395506; O15397:IPO8; NbExp=3; IntAct=EBI-528269, EBI-358808; Q9UHD2:TBK1; NbExp=2; IntAct=EBI-528269, EBI-356402; A7MCY6:TBKBP1; NbExp=2; IntAct=EBI-528269, EBI-359969; SUBCELLULAR LOCATION: Cytoplasm, P-body. Nucleus. Note=Translational repression of mRNAs results in their recruitment to P-bodies. Translocation to the nucleus requires IMP8. DOMAIN: The Piwi domain may perform RNA cleavage by a mechanism similar to that of RNase H. However, while RNase H utilizes a triad of Asp-Asp-Glu (DDE) for metal ion coordination, this protein appears to utilize a triad of Asp-Asp-His (DDH). PTM: Hydroxylated. 4-hydroxylation appears to enhance protein stability but is not required for miRNA-binding or endonuclease activity. SIMILARITY: Belongs to the argonaute family. Ago subfamily. SIMILARITY: Contains 1 PAZ domain. SIMILARITY: Contains 1 Piwi domain. SEQUENCE CAUTION: Sequence=AAH07633.1; Type=Erroneous initiation; Sequence=AAL76093.1; Type=Miscellaneous discrepancy; Note=cDNA contains a duplication of an internal sequence at the 5' end; Sequence=BC125214; Type=Frameshift; Positions=450;
The RNAfold program from the Vienna RNA Package is used to perform the secondary structure predictions and folding calculations. The estimated folding energy is in kcal/mol. The more negative the energy, the more secondary structure the RNA is likely to have.
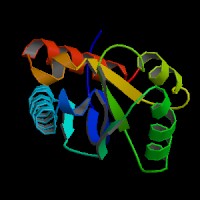
ModBase Predicted Comparative 3D Structure on Q9UKV8
Front
Top
Side
The pictures above may be empty if there is no ModBase structure for the protein. The ModBase structure frequently covers just a fragment of the protein. You may be asked to log onto ModBase the first time you click on the pictures. It is simplest after logging in to just click on the picture again to get to the specific info on that model.
Orthologous Genes in Other Species
Orthologies between human, mouse, and rat are computed by taking the best BLASTP hit, and filtering out non-syntenic hits. For more distant species reciprocal-best BLASTP hits are used. Note that the absence of an ortholog in the table below may reflect incomplete annotations in the other species rather than a true absence of the orthologous gene.
 Sequence and Links to Tools and Databases
Sequence and Links to Tools and Databases  Common Gene Haplotype Alleles
Common Gene Haplotype Alleles