ID:DAG1_HUMAN DESCRIPTION: RecName: Full=Dystroglycan; AltName: Full=Dystrophin-associated glycoprotein 1; Contains: RecName: Full=Alpha-dystroglycan; Short=Alpha-DG; Contains: RecName: Full=Beta-dystroglycan; Short=Beta-DG; Flags: Precursor; FUNCTION: The dystroglycan complex is involved in a number of processes including laminin and basement membrane assembly, sarcolemmal stability, cell survival, peripheral nerve myelination, nodal structure, cell migration, and epithelial polarization. FUNCTION: Alpha-dystroglycan is an extracellular peripheral glycoprotein that acts as a receptor for both extracellular matrix proteins containing laminin-G domains, and for certain adenoviruses. Receptor for laminin-2 (LAMA2) and agrin in peripheral nerve Schwann cells. Also acts as a receptor for M.leprae in peripheral nerve Schwann cells but only in the presence of the G-domain of LAMA2, and for lymphocytic choriomeningitis virus, Old World Lassa fever virus, and clade C New World arenaviruses. FUNCTION: Beta-dystroglycan is a transmembrane protein that plays important roles in connecting the extracellular matrix to the cytoskeleton. Acts as a cell adhesion receptor in both muscle and non-muscle tissues. Receptor for both DMD and UTRN and, through these interactions, scaffolds axin to the cytoskeleton. Also functions in cell adhesion-mediated signaling and implicated in cell polarity. SUBUNIT: Monomer. Heterodimer of alpha- and beta-dystroglycan subunits which are the central components of the dystrophin- glycoprotein complex. This complex then can form a dystrophin- associated glycoprotein complex (DGC) which is composed of three subcomplexes: a cytoplasmic complex comprised of DMD (or UTRN), DTNA and a number of syntrophins, such as SNTB1, SNTB2, SNTG1 and SNTG2, the transmembrane dystroglycan complex, and the sarcoglycan-sarcospan complex. Interacts (via the N-terminal of alphaDAG1) with LARGE; the interaction enhances laminin binding (By similarity). Interacts with SGCD. Interacts with AGR2 and AGR3. Interacts (betaDAG1) with DMD; the interaction is inhibited by phosphorylaion on the PPXY motif. Interacts (betaDAG1, via its PPXY motif) with UTRN (via its WWW and ZZ domains); the interaction is inhibited by phosphorylation on the PPXY motif. Interacts (betaDAG1, via its phosphorylated PPXY motif) with the SH2 domain-containing proteins, FYN, CSK, NCK and SHC. Interacts (betaDAG1) with CAV3 (via a central WW-like domain); the interaction disrupts the binding of DMD. BetaDAG1 directly interacts with ANK3, but not with ANK2; this interaction does not interfere with DMD-binding and is required for retention at costameres (By similarity). INTERACTION: Q3T1J5:Ank3 (xeno); NbExp=2; IntAct=EBI-1755945, EBI-2133962; P16333:NCK1; NbExp=2; IntAct=EBI-1755945, EBI-389883; SUBCELLULAR LOCATION: Alpha-dystroglycan: Secreted, extracellular space. SUBCELLULAR LOCATION: Beta-dystroglycan: Cell membrane; Single- pass type I membrane protein. Cytoplasm, cytoskeleton. Nucleus, nucleoplasm. Cell membrane, sarcolemma (By similarity). Cell junction, synapse, postsynaptic cell membrane (By similarity). Note=The monomeric form translocates to the nucleus via the action of importins and depends on RAN. Nuclear transport is inhibited by Tyr-892 phosphorylation. In skeletal muscle, this phosphorylated form locates to a vesicular internal membrane compartment. In muscle cells, sarcolemma localization requires the presence of ANK2, while localization to costameres requires the presence of ANK3. Localizes to neuromuscular junctions (NMJs) in the presence of ANK2 (By similarity). In peripheral nerves, localizes to the Schwann cell membrane. Colocalizes with ERM proteins in Schwann- cell microvilli. TISSUE SPECIFICITY: Expressed in a variety of fetal and adult tissues. In epidermal tissue, located to the basement membrane. Also expressed in keratinocytes and fibroblasts. PTM: O- and N-glycosylated. Alpha-dystroglycan is heavily O- glycosylated comprising of up to two thirds of its mass and the carbohydrate composition differs depending on tissue type. Mucin- type O-glycosylation is important for ligand binding activity. O- mannosylation of alpha-DAG1 is found in high abundance in both brain and muscle where the most abundant glycan is Sia-alpha-2-3- Gal-beta-1-4-Glc-NAc-beta-1-2-Man. In muscle, glycosylation on Thr-379 by a phosphorylated O-mannosyl glycan with the structure 2-(N-acetylamido)-2-deoxygalactosyl-beta-1,3-2-(N-acetylamido)-2- deoxyglucosyl-beta-1,4-6-phosphomannose is mediated by like- acetylglucosaminyltransferase (LARGE) protein and is required for laminin binding. O-mannosylation is also required for binding lymphocytic choriomeningitis virus, Old World Lassa fever virus, and clade C New World arenaviruses. The O-glycosyl hexose on Thr- 367, Thr-369, Thr-372, Thr-381 and Thr-388 is probably mannose. O- glycosylated in the N-terminal region with a core 1 or possibly core 8 glycan. The beta subunit is N-glycosylated. PTM: Autolytic cleavage produces the alpha and beta subunits. In cutaneous cells, as well as in certain pathological conditions, shedding of beta-dystroglcan can occur releasing a peptide of about 30 kDa. PTM: SRC-mediated phosphorylation of the PPXY motif of the beta subunit recruits SH2 domain-containing proteins, but inhibits binding to WWW domain-containing proteins, DMD and UTRN. This phosphorylation also inhibits nuclear entry. DISEASE: Defects in DAG1 are the cause of muscular dystrophy- dystroglycanopathy limb-girdle type C7 (MDDGC7) [MIM:613818]. An autosomal recessive muscular dystrophy showing onset in early childhood, and associated with mental retardation without structural brain anomalies. Note=MDDGC7 is caused by DAG1 mutations that interfere with normal post-translational processing, resulting in defective DAG1 glycosylation and impaired interactions with extracellular-matrix components. Other muscular dystrophy-dystroglycanopathies are caused by defects in enzymes involved in protein O-glycosylation. SIMILARITY: Contains 1 peptidase S72 domain. WEB RESOURCE: Name=Wikipedia; Note=Dystroglycan entry; URL="http://en.wikipedia.org/wiki/Dystroglycan";
The RNAfold program from the Vienna RNA Package is used to perform the secondary structure predictions and folding calculations. The estimated folding energy is in kcal/mol. The more negative the energy, the more secondary structure the RNA is likely to have.
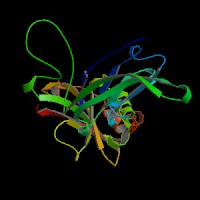
ModBase Predicted Comparative 3D Structure on Q14118
Front
Top
Side
The pictures above may be empty if there is no ModBase structure for the protein. The ModBase structure frequently covers just a fragment of the protein. You may be asked to log onto ModBase the first time you click on the pictures. It is simplest after logging in to just click on the picture again to get to the specific info on that model.
Orthologous Genes in Other Species
Orthologies between human, mouse, and rat are computed by taking the best BLASTP hit, and filtering out non-syntenic hits. For more distant species reciprocal-best BLASTP hits are used. Note that the absence of an ortholog in the table below may reflect incomplete annotations in the other species rather than a true absence of the orthologous gene.
 Sequence and Links to Tools and Databases
Sequence and Links to Tools and Databases  Common Gene Haplotype Alleles
Common Gene Haplotype Alleles